
The University of Minnesota’s Dr Andres Gomez, Dr Isaac Salfer, and Dr Marshall Stern discuss how the rumen microbiome is influenced by non-nutritional factors, affecting ruminant livestock performance.
The vast array of microorganisms that inhabit the forestomach of ruminants – mainly bacteria, fungi, virus, and protozoa (also known as the rumen microbiome) – play an indispensable role in determining how the animal retrieves energy from feed, and hence profoundly affects animal productive performance.
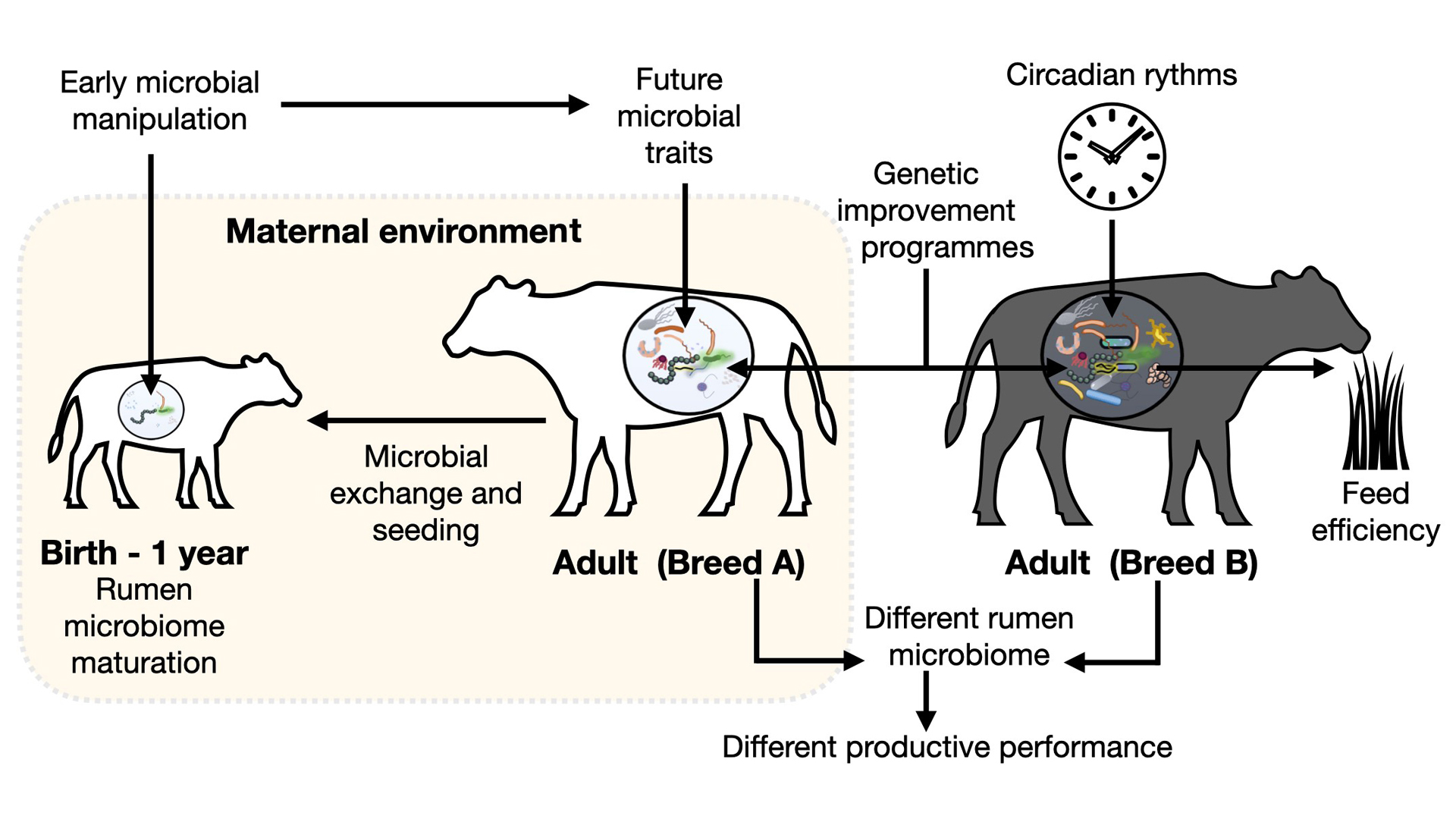
A significant body of research has demonstrated how the nutritional composition of diets modulate the rumen microbiome, impacting animal production. However, there are important factors, beyond diet and nutrition, that also affect the rumen microbiome, and hence animal production. Among these factors, early life and the maternal environment are important modulators of the primordial microbiome acquired by ruminants, with impacts on health and productive performance that could extend to later in life.
It is for this reason that recent research has focused on elucidating strategies to influence the way microbes first colonise the animal rumen to determine desirable microbiome and physiological traits along the life course.
Another non-dietary factor that significantly impacts rumen microbial communities is the genetic background of the animal. Consequently, animal genetic improvement programmes have been recently proposed to not only select for desirable productive and phenotypic traits, but also to pick specific microbial species in the rumen that play a role in impacting productive efficiency and performance.
Other important non-nutritional factors that affect the rumen microbiome have been shown to operate on a narrower scale, such as time of day. Pioneering research has shown that circadian rhythms can affect productive performance, and that the rumen microbiome may have a major influence on this process. In this article, we present an overview of how non-nutritional factors impact the rumen microbiome in dairy and beef livestock and discuss how these relationships need to be considered to improve the efficiency of ruminant production systems.
Impact of early life on the rumen microbiome
The first microbiome in animals is acquired at birth. During the birthing process, microbes in the mother’s vaginal canal, oral cavity, colostrum, faeces, skin and microbes from the birthing environment promptly colonise and seed the newborn’s body.1,2,3 In ruminants, it has been shown that, although the early rumen is largely underdeveloped and the intestine and abomasum serve the main digestive functions, it harbours bacteria active in degrading and fermenting fibrous feedstuffs as early as one week of age, even in the absence of a solid diet.4,5 This process is essential for the maturation of the rumen; because, without this colonisation, ruminants would be unable to digest dietary fibre as a source of energy. The main reason is that, as ruminants transition to forage-based diets, they strictly depend on fibre degradation in the rumen for extracting energy from feed.
Colonisation of the rumen is first done by bacteria, followed by an increase in the diversity of archaea, fungi, and protozoa during the first two weeks of life.6 This early colonisation process may continue for a year until the rumen microbiome is functionally and structurally developed.7
In addition to providing the ability to metabolise feed, microbial colonisation is instrumental in the maturation of the calf’s gut-associated immune system.8,9 Therefore, the early microbial colonisation process in the rumen not only impacts how animals extract energy from feed, but also how they respond to infectious disease challenges. Furthermore, the establishment of the first microbes, the fermentative functions performed by rumen microorganisms, and the nature of pre-and post-weaning diets, contributes to the anatomical and functional maturation of the rumen as an organ, including growth of rumen absorptive surfaces and the musculature responsible for rumen contractions.10
Early management decisions related to lactation or background diets, as well as birthing environments, may impact the way microbes first colonise the young rumen. For example, the environment in
which beef calves are reared (e.g., two different farms) affects the composition and temporal dynamics of bacterial and archaeal colonisers in the early rumen.11
The age at which dairy calves are weaned can also affect early establishment of rumen microbes; for instance, it has been shown that, when Holstein calves are weaned early (seven weeks of age), the rumen microbiome is able to abruptly adapt to the challenges brought by a change to solid, fibrous feed through key functions performed by fibre degraders. In contrast, during late weaning at 17 weeks of age, the rumen microbiome seems more resilient and less affected by a change to solid feed.12
Research with lambs demonstrated that exposure to maternal sources of colostrum, compared with colostrum replacer, preserves communities of fibrolytic protozoa and fungi beyond weaning, which translated to higher feed digestibility and growth during the grazing period.13 This body of work demonstrated that the early rearing environment, including the degree of maternal contact and microbial exposures, could determine the rate at which the rumen microbiome matures, and how microbes with important metabolic roles, such as fibre and starch degradation or methanogenic archaea, colonise the rumen.
Early life management interventions have emerged as key opportunities for shaping rumen microbiomes and achieving desirable phenotypes. Exposure to or inoculation of young calves with ruminal fluid of adult animals promises to be a potential strategy to influence the rumen microbiome at an early age. However, the effect of this intervention appears to be more influential on animal health than performance, with research demonstrating a reduction in post-weaning diarrhoea incidence but minimal effects on growth rate.14,15 This inoculation procedure is analogous to what faecal material transplants (FMT) can achieve in humans, showing that this procedure can ameliorate the detrimental effects of digestive disorders such as C. difficile infections and inflammatory bowel diseases.16 Exposure of young calves to rumen fluid of adult individuals could potentially be used to strengthen calf health along the life course.
The impact of genetics
Individual animal genetics impact the composition of the rumen microbiome. Assessing the genetic impacts on microbial community composition is challenging due to the impact of diet and environmental factors. However, recent experiments elucidated some effects.
Roehe et al. (2016) observed that different sire progeny groups (groups of offspring of a single bull) differed greatly in the amount of methane they produced.17 They found that differences were largely due to microbial genes associated with methane production, and with genes involved in interactions between the animal and its microbiome. Similarly, Difford et al. (2018) detected moderate genetic influence on the communities of rumen bacteria and methanogenic archaea.18 As a reference point, milk production is typically considered a highly heritable trait.19
Breed also impacts composition of the microbiome between different livestock species. Li et al. (2016) observed differences in the diversity of rumen microbes between Angus, Charolais and Angus x Charolais beef cow crosses.20 Furthermore, Noel et al. (2019) detected differences in the kind of microbial species in the rumen, particularly different methanogenic microbes between Holstein and Jersey dairy cow breeds.21 However, they were unable to find any significant associations between total methane emissions. Finally, they noticed that the microbiome of Jersey cows is more resistant to changes in diet than that of Holsteins.
Additionally, in crossbred beef cattle, there is an influence of the sire breed on the rumen microbiome of the progeny.22 However, our preliminary data has shown that, when feed is kept the same, there are no distinguishable differences in the rumen microbiome between crossbred cows (Holstein x Viking Red x Montbe éliarde, also known as ProCROSS) and purebred Holsteins. This is a remarkable finding given that ProCROSS-bred cows show lower dry matter intake compared with pure Holstein cows.23 Factors other than the rumen microbiome indirectly impact rumen microbial communities, and drive differences in productive efficiency between animals of different genetic backgrounds.
One factor responsible for genetic differences in microbiome composition may be related to differences in the rates of nutrient flow out of the rumen. Nutrient flow rates can impact the washout and growth of certain bacterial and protozoal species.24
Diet is the primary driver of rumen nutrient flow rates, but it is also impacted by differences in feeding behaviour and anatomical differences such as rumen size. Smuts et al. (1995) determined that there is a genetic component to rumen passage rates, showing high heritability of this physiological trait in Merino rams.25 Li et al. (2016) found associations between decreased feeding frequency and variations in the rumen microbiota between Angus x Charolais crosses and purebred Angus or Charolais cattle.
Currently, none of the major livestock species directly select for traits such as individual microbial population or methane production. However, a trait called residual feed intake (RFI), which is an indicator of feed efficiency, was recently developed as a trait for genetic selection of dairy cattle.26
The heritability for RFI is low.27 It is likely that the genetic variation in this trait is partially due to differences in the rumen microbiome. Future genetic improvement programmes may consider directly selecting for characteristics of the rumen microbiome, such as diversity or abundance of methanogenic archaea, to give perhaps a more precise and heritable way to improve feed efficiency and the carbon footprint of ruminant production.
Pioneering research on elucidating heritability of rumen microbiome species has shown that microbes with high heritability scores are associated with key metabolic traits, such as ratios of volatile fatty acids produced, methane metabolism and concentration of amino acids in the rumen.28 Moreover, the abundance of highly heritable rumen taxa is also associated with dry matter intake, feed efficiency and milk fat composition29 (Li et al., 2019), although these associations seem to be of lesser magnitude than those found with specific rumen metabolic traits.
The main characteristic of microbes with high heritability scores in the rumen is their involvement in fibre metabolism and production of volatile fatty acids (e.g. Prevotella, Fibrobacteres and Neocallimastix).30 Despite the fact that values of microbial heritability can be highly variable across different studies, information of heritability of specific rumen bacteria, fungi and protozoa can be used to predict the efficiency of beef and dairy production systems at an early stage.
The impact of the time of day
Daily changes in the rumen microbiome are well characterised. Bryant and Robinson (1961) reported that total bacterial colony counts in the rumen differed across the day, and that the daily pattern of bacterial abundance differed by type of diet.31
They observed that, with both diets, bacterial counts increased rapidly after feeding, but a grain-based diet resulted in a sharp decline approximately six hours after feeding, whereas a hay-based diet resulted in high bacterial counts persisting for nearly 20 hours, before declining just before the next day’s feeding. The bacteriophage population of the rumen also displays a waveform pattern that peaks approximately eight to ten hours after feeding.32
One factor that influences daily changes in the rumen microbiome is the circadian rhythms of the animal. Circadian rhythms are repeating cycles of physiology and behaviour that repeat over 24 hours. A key component of circadian rhythms is that they are endogenous, meaning that they are generated by an internal mechanism within an organism’s cells, and they persist in constant environmental conditions. In mammals, they are regulated by a core set of ‘clock’ transcription factors that regulate the gene expression over a 24-hour period.33 Many important cellular processes related to growth, metabolism, and reproduction are regulated by circadian rhythms.
The feeding pattern of mammals is regulated directly by ‘food-entrainable circadian oscillators’ that coordinate signals from the light-dark cycle and food intake.34 Most ruminant animals are naturally crepuscular, meaning that they eat the majority of their food at dawn and dusk, with very minimal feed intake overnight. However, in livestock production systems, management can have major impacts on feeding patterns. For dairy cows, feed intake increases when fresh feed is delivered to cows, and when cows have finished being milked.
The daily pattern of intake influences the amount of feed entering the rumen across the day, with three to eight times more fermentable substrate entering the rumen during high-intake periods of the day.35 This can subsequently impact the pH of the rumen, with pH typically peaking before feeding and reaching a minimum ten hours later.36 In addition to the feeding pattern, host circadian rhythms affect the daily pattern of rumination. Rumination is a true circadian rhythm that has been shown to be free under constant lighting.37 While it is not entrained by light, the circadian rhythm of rumination is affected by feeding time, and typically follows a daily pattern that is inverse of feed intake.38
Another factor that is not well-characterised but could potentially influence the daily patterns of rumen microbial abundance is circadian rhythms within the microbes themselves. Historically, prokaryotic microorganisms were considered to be too ‘simplistic’ and too short-lived to have evolved the capacity of having circadian rhythms. However, Grobbelaar et al. (1986) discovered that the ability to fixate nitrogen by the cyanobacterial species Synechococcus elongatus followed a circadian rhythm that persisted in constant environmental conditions.39
It was later discovered these rhythms are generated by a phosphorylation-based circadian mechanism that is entrained by light.40 It is currently unknown if a similar mechanism occurs in the anaerobic bacteria of the rumen. Yerushalmi and Volesky (1989) detected 24-hour cyclic patterns of anaerobic fermentation and short chain fatty acid production in pure continuous cultures of Clostridium acetobutylicum, suggesting that there is potential for further exploration into the circadian rhythms of rumen microbes.41
Our research group has begun to conduct research investigating the diurnal patterns of rumen microbial populations. We have observed that several key rumen bacterial species follow daily patterns that are influenced by feeding time. We conducted an experiment where we fed cows three different feeding regimens: a high-fibre diet in the morning and a high-starch diet in the afternoon (High/Low); a high-starch diet in the morning and a high-fibre diet in the evening (Low/High); and a moderate starch and fibre fed 1x/d in the morning (CON).42
We observed that the feeding regimen was found to have a large impact on the daily pattern of certain microbial species. Streptococcus bovis, a species with a known role in the development of rumen acidosis, peaked in the High/Low and Low/High feeding regimens, but not the CON treatment. Recent unpublished data from our group suggested that microbial diversity within the rumen varies by time of day and diet, peaking near the middle of the day in both high-starch and low-starch diets, with a greater peak occurring when high starch diets are fed.
Conclusion
Modulation of the rumen microbiome, beyond nutritional strategies, opens a window of opportunity for improving animal health and productivity (Fig. 1). The immature rumen microbial environment that calves display at an early age can be readily modulated to imprint desirable metabolic and health characteristics that can extend into subsequent stages of development. Likewise, the realisation that rumen microbiome traits are heritable and tightly associated with the genetic background or breed of the animal, suggests that genetic improvement programmes need to move from only selecting for specific genetic or phenotypic traits to also include the microbiome.
Lastly, the emergence of circadian biology research in animal production offers new perspectives on management strategies that can ensure that animals utilise feed in the most efficient ways. As the field continues to leverage nucleic acid sequencing techniques and in-vitro mechanistic models to elucidate the composition and role of microbes in the animal rumen in the context of age, genetics and circadian biology, the manipulation of the rumen environment to improve the efficiency of animal production is close to becoming a reality for small and large-scale production systems.
References
1 Dominguez-Bello, M.G. et al. (2010) ‘Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns’, Proceedings of the National Academy
of Sciences of the United States of America, 107(26), pp. 11971–11975
2 Yeoman, C.J. et al. (2018) ‘Biogeographical Differences in the Influence of Maternal Microbial Sources on the Early Successional Development of the Bovine Neonatal Gastrointestinal tract’, Scientific Reports, 8(1), p. 3197
3 Klein-Jöbstl, D. et al. (2019) ‘Microbiota of newborn calves and their mothers reveals possible transfer routes for newborn calves’ gastrointestinal microbiota’, PloS one, 14(8), p. e0220554
4 Castro, J.J. et al. (2016) ‘Changes in the intestinal bacterial community, short-chain fatty acid profile, and intestinal development of preweaned Holstein calves. 2. Effects of gastrointestinal site and age’, Journal of Dairy Science, 99(12), pp. 9703–9715
5 Malmuthuge, N., Liang, G. and Guan, L.L. (2019) ‘Regulation of rumen development in neonatal ruminants through microbial metagenomes and host transcriptomes’, Genome Biology, 20(1),
p. 172
6 Minato, H. et al. (1992) ‘COLONIZATION OF MICROORGANISMS IN THE RUMEN OF YOUNG CALVES’, The Journal of General and Applied Microbiology, 38(5), pp. 447–456
7 Jami, E. et al. (2013) ‘Exploring the bovine rumen bacterial community from birth to adulthood’, The ISME Journal, 7(6), pp. 1069–1079
8 Taschuk, R. and Griebel, P.J. (2012) ‘Commensal microbiome effects on mucosal immune system development in the ruminant gastrointestinal tract’, Animal Health Research Reviews / Conference of Research Workers in Animal Diseases, 13(1), pp. 129–141
9 Chase, C. and Kaushik, R.S. (2019) ‘Mucosal Immune System of Cattle: All Immune Responses Begin Here’, The Veterinary Clinics of North America: Food Animal Practice, 35(3), pp. 431–451
10 Yáñez-Ruiz, D.R., Abecia, L. and Newbold, C.J. (2015) ‘Manipulating rumen microbiome and fermentation through interventions during early life: a review’, Frontiers in Microbiology, 6, p. 1133
11 O’Hara, E. et al. (2020) ‘Investigating temporal microbial dynamics in the rumen of beef calves raised on two farms during early life’, FEMS Microbiology Ecology, 96(2) doi:10.1093/femsec/fiz203
12 Amin, N. et al. (2021) ‘Evolution of rumen and oral microbiota in calves is influenced by age and time of weaning’, Animal Microbiome, 3(1), p. 31
13 Belanche, A. et al. (2019) ‘Maternal versus artificial rearing shapes the rumen microbiome having minor long-term physiological implications’, Environmental Microbiology, 21(11), pp. 4360–4377
14 Bu, D. et al. (2020) ‘Repeated Inoculation of Young Calves With Rumen Microbiota Does Not Significantly Modulate the Rumen Prokaryotic Microbiota Consistently but Decreases Diarrhea’, Frontiers in Microbiology, 11, p. 1403
15 Park, T. et al. (2021) ‘Pre-weaning Ruminal Administration of Differentially-Enriched, Rumen-Derived Inocula Shaped Rumen Bacterial Communities and Co-occurrence Networks of Post-weaned Dairy Calves’, Frontiers in Microbiology, 12, p. 625488
16 Khoruts, A. and Sadowsky, M.J. (2016) ‘Understanding the mechanisms of faecal microbiota transplantation’, Nature Reviews. Gastroenterology & Hepatology, 13(9), pp. 508–516
17 Roehe, R. et al. (2016) ‘Bovine Host Genetic Variation Influences Rumen Microbial Methane Production with Best Selection Criterion for Low Methane Emitting and Efficiently Feed Converting Hosts Based on Metagenomic Gene Abundance’, PLoS Genetics, 12(2), p. e1005846
18 Difford, G.F. et al. (2018) ‘Host genetics and the rumen microbiome jointly associate with methane emissions in dairy cows’, PLoS Genetics, 14(10), p. e1007580
19 Elgersma et al. (2018) Fluctuations in milk yield are heritable and can be used as a resilience indicator to breed healthy cows. Dairy Science
20 Li, Z. et al. (2016) ‘Changes in the rumen microbiome and metabolites reveal the effect of host genetics on hybrid crosses’, Environmental Microbiology Reports, 8(6), pp. 1016–1023
21 Noel, S.J. et al. (2019) ‘Rumen and Fecal Microbial Community Structure of Holstein and Jersey Dairy Cows as Affected by Breed, Diet, and Residual Feed Intake’, Animals : An Open Access Journal from MDPI, 9(8). doi:10.3390/ani9080498
22 Hernandez-Sanabria, E. et al. (2013) ‘Influence of sire breed on the interplay among rumen microbial populations inhabiting the rumen liquid of the progeny in beef cattle’, PloS One, 8(3), p. e58461
23 Shonka-Martin, B.N. (2018) Comparison of ProCROSS and Holstein Cows for Dry Matter Intake, Body Weight, Cow Height, Body Condition Score, Production, Feed Efficiency, Income over Feed Cost, and Residual Feed Intake. Edited by L.B. Hansen and B.J. Heins. University of Minnesota.
24 Stern, M.D., Ortega, M.E. and Satter, L.D. (1983) ‘Retention time in rumen and degradation of protein supplements fed to lactating dairy cattle’, Journal of Dairy Science, 66(6), pp. 1264–1271
25 Smuts, M., Meissner, H.H. and Cronjé, P.B. (1995) ‘Retention time of digesta in the rumen: its repeatability and relationship with wool production of Merino rams’, Journal of Animal Science, 73(1), pp. 206–210
26 Connor, E.E. et al. (2013) ‘Use of residual feed intake in Holsteins during early lactation shows potential to improve feed efficiency through genetic selection’, Journal of Animal Science, 91(8), pp. 3978–3988
27 VandeHaar, M.J. et al. (2016) ‘Harnessing the genetics of the modern dairy cow to continue improvements in feed efficiency’, Journal of Dairy Science, 99(6), pp. 4941–4954
28 Sasson, G. et al. (2017) ‘Heritable Bovine Rumen Bacteria Are Phylogenetically Related and Correlated with the Cow’s Capacity To Harvest Energy from Its Feed’, mBio. doi:10.1128/mbio.00703-17
29 Li, F. et al. (2019) ‘Host genetics influence the rumen microbiota and heritable rumen microbial features associate with feed efficiency in cattle’, Microbiome, 7(1), p. 92
30 Wallace, R.J. et al. (2019) ‘A heritable subset of the core rumen microbiome dictates dairy cow productivity and emissions’, Science Advances, 5(7), p. eaav8391
31 Bryant, M.P. and Robinson, I.M. (1961) ‘An Improved Nonselective Culture Medium for Ruminal Bacteria and Its Use in Determining Diurnal Variation in Numbers of Bacteria in the Rumen’, Journal of Dairy Science, 44(8), pp. 1446–1456
32 Klieve, A.V. and Swain, R.A. (1993) ‘Estimation of ruminal bacteriophage numbers by pulsed-field gel electrophoresis and laser densitometry’, Applied and Environmental Microbiology, 59(7), pp. 2299–2303
33 Cox, K.H. and Takahashi, J.S. (2019) ‘Circadian clock genes and the transcriptional architecture of the clock mechanism’, Journal of Molecular Endocrinology, 63(4), pp. R93–R102
34 Stephan, F.K. (2002) ‘The ‘Other’ Circadian System: Food as a Zeitgeber’, Journal of Biological Rhythms, 17(4), pp. 284–292
35 Ying, Y. et al. (2015) ‘The effects of feeding rations that differ in neutral detergent fibre and starch concentration within a day on rumen digesta nutrient concentration, pH, and fermentation products in dairy cows’, Journal of Dairy Science, 98(7), pp. 4685–4697
36 Yang, W.Z. and Beauchemin, K.A. (2006) ‘Effects of Physically Effective Fibre on Chewing Activity and Ruminal pH of Dairy Cows Fed Diets Based on Barley Silage’, Journal of Dairy Science, pp. 217–228. doi:10.3168/jds.s0022-0302(06)72086-0
37 Gordon, J.G. and McAllister, I.K. (1970) ‘The circadian rhythm of rumination’, The Journal of Agricultural Science, 74(2), pp. 291–297
38 Salfer, I.J. et al. (2018) ‘The effects of source and concentration of dietary fibre, starch, and fatty acids on the daily patterns of feed intake, rumination, and rumen pH in dairy cows’, Journal of Dairy Science, 101(12), pp. 10911–10921
39 Grobbelaar, N. et al. (1986) ‘Dinitrogen-fixing endogenous rhythm in IRF-1’, FEMS Microbiology Letters, 37(2), pp. 173–177
40 Swan, J.A. et al. (2018) ‘Structure, function, and mechanism of the core circadian clock in cyanobacteria’, The Journal of Biological Chemistry, 293(14), pp. 5026–5034
41 Yerushalmi, L. et al. (1989) ‘Circadian rhythmicity in a fermentation process?’, Applied Microbiology and Biotechnology, 30(5). doi:10.1007/bf00263850
42 Salfer, I.J. et al. (2021) ‘The effects of feeding rations that differ in neutral detergent fibre and starch within a day on the daily pattern of key rumen microbial populations’, JDS Communications, 2(6), pp. 334–339
Please note, this article will also appear in the ninth edition of our quarterly publication





